- 品牌
- 有梦生物
- 服务项目
- 扫描
0. 全景扫描助力**研究,对**组织切片进行全域扫描时,可同时识别*细胞的空间分布模式、增殖活性及基因突变类型,结合基因测序数据中的突变位点与表达谱,能深入分析**微环境中免疫细胞的浸润程度、*细胞的血管新生情况及二者的相互作用机制。它为精细*****方案制定提供**全景病理信息,例如在肺****中,通过确定****区域与边缘区域的差异特征,可指导个性化放疗方案的制定,提高***效果并减少对正常组织的损伤,同时为新型免疫***药物的疗效评估提供直观依据。全景扫描追踪精子获能过程,记录其穿越透明带的关键形态变化。贵州免疫组化全景扫描单价
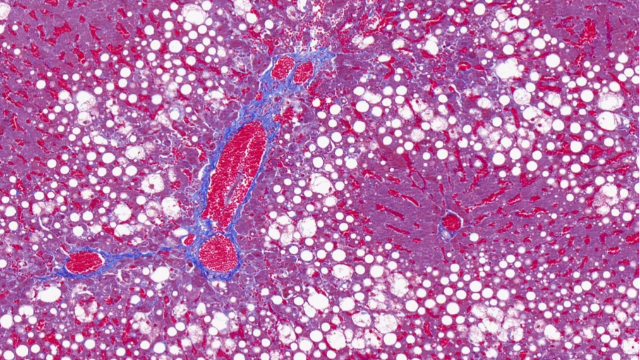
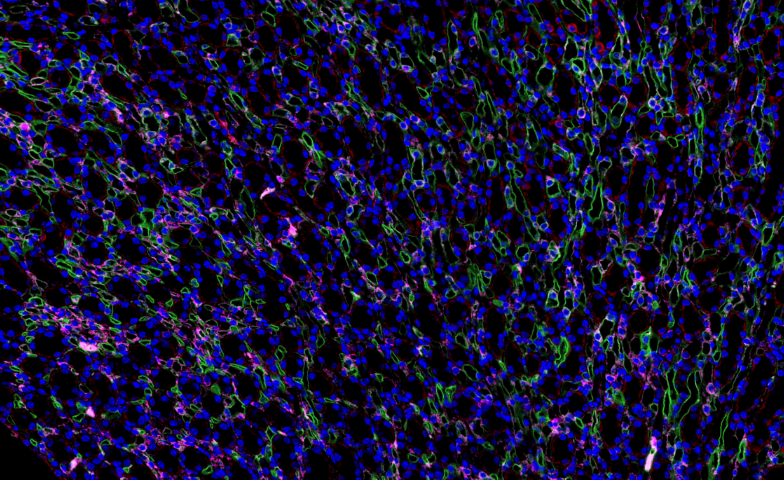
全景扫描在动物行为学研究中用于记录动物的整体行为模式及与环境的互动,通过红外摄像与运动捕捉技术结合,对动物的觅食、交配、社群互动等行为进行全景拍摄与分析,提取行为参数如活动范围、运动速度、互动频率等。结合神经影像学数据,揭示行为背后的神经机制,例如在研究小鼠的焦虑行为时,全景扫描发现了小鼠在旷场实验中的活动轨迹与大脑特定区域神经元活动的关联,为理解焦虑症的神经基础提供了线索,也为抗焦虑药物的筛选提供了行为学评估方法。贵州免疫组化全景扫描单价对红树林根系全景扫描,探究其在潮间带的固着与通气适应机制。

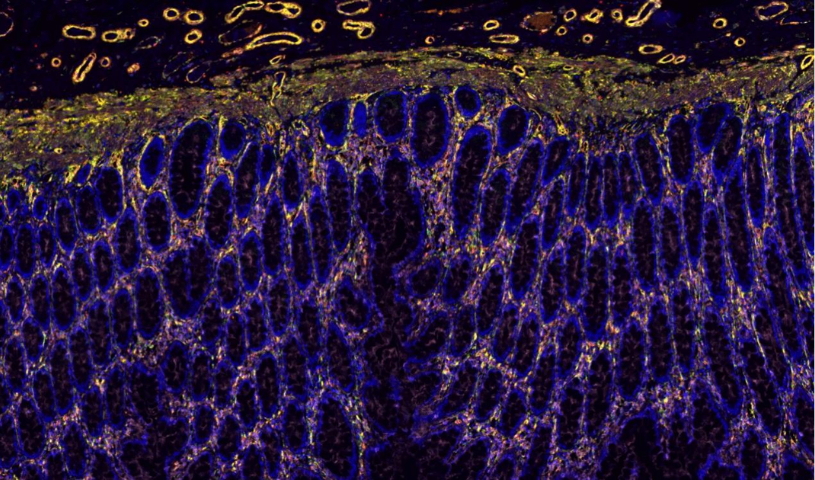
在植物光合作用研究中,全景扫描技术 通过多尺度成像与功能分析联用,系统揭示了 光合结构-功能耦合机制。该技术整合 冷冻电镜断层扫描(Cryo-ET)、荧光寿命成像(FLIM)和 原子力显微镜(AFM),实现了从 类囊体基粒堆叠(单层厚度10-12nm)到 全叶光合活性 的跨维度解析。以高光胁迫(1500μmol·m⁻²·s⁻¹)研究为例:超微结构层面:冷冻电镜全景扫描 显示PSII超复合体在强光下2小时内发生 二聚体解离(从80%降至35%)类囊体膜出现穿孔(直径50-100nm),伴随 Cyt b6f复合体空间重排生理动态层面:多光谱荧光扫描 捕获到叶黄素循环(VDE酶***)在5分钟内启动,非光化学淬灭(NPQ)效率提升3倍拉曼成像 发现β-胡萝卜素在强光区优先降解(1530cm⁻¹特征峰减弱60%)分子调控层面:原位杂交全景扫描 显示 PsbS基因 在束鞘细胞中表达量激增8倍,与抗光氧化关键蛋白(如PTOX)共定位
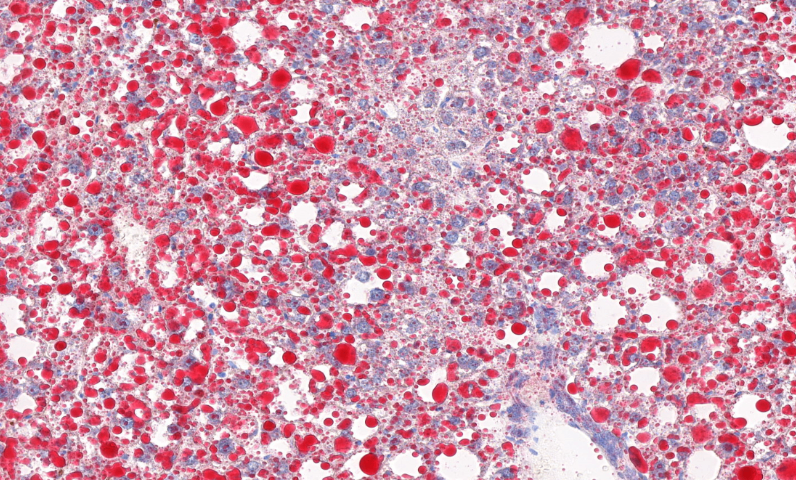
结合稳定同位素示踪技术,全景扫描进一步阐明了土壤团聚体 对碳封存的影响:微团聚体(<250μm)通过物理保护作用减缓有机碳的微生物降解,而大团聚体的形成则依赖于***菌丝和根系分泌物的胶结作用。这些发现为可持续农业 提供了重要依据,例如通过调整耕作方式优化孔隙结构,或接种特定微生物群落增强土壤肥力。此外,在污染土壤修复 领域,全景扫描揭示了污染物(如重金属、微塑料)在孔隙中的迁移规律,为开发靶向生物修复 策略奠定了基础。未来,结合人工智能图像分析,该技术有望在土壤碳汇评估和气候变化应对中发挥更大作用。全景扫描监测病毒出芽释放,展示子代病毒从宿主细胞脱离的过程。
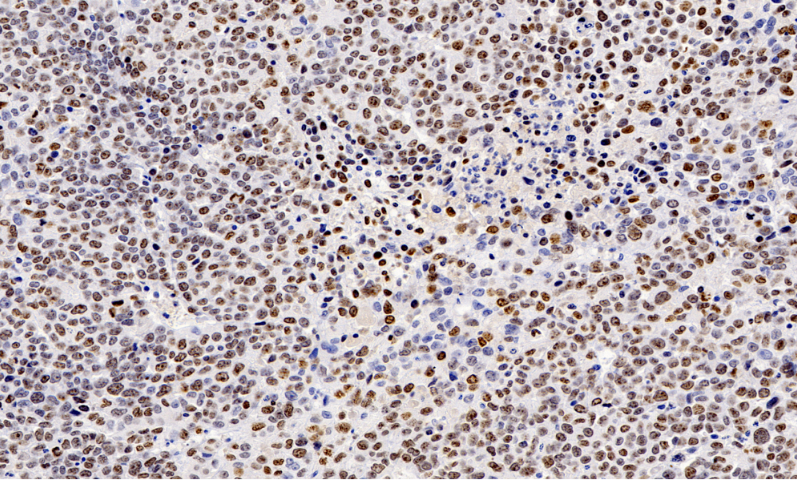
这些发现直接指导了光合增效工程:通过CRISPR编辑LHCII磷酸化位点,使水稻在强光下维持90%以上的Fv/Fm值。***研发的纳米探针标记技术,可实时监测单个叶绿体质子动力势(ΔpH)变化,为开发"智能光保护"作物提供了新工具。该技术已成功应用于C4植物进化研究,通过全景扫描玉米花环结构,揭示叶肉细胞-维管束鞘细胞间的代谢物通道密度与CO2浓缩效率呈正相关(R²=0.92)。这些突破不仅阐明了光合机构的损伤修复机制,更为设计新一代光合生物反应器提供了结构仿生模板。全景扫描监测*细胞转移,追踪其在血管内的移动及侵袭组织过程。贵州免疫组化全景扫描单价
全景扫描追踪根系分泌物,记录其在根际土壤中的扩散与作用范围。贵州免疫组化全景扫描单价
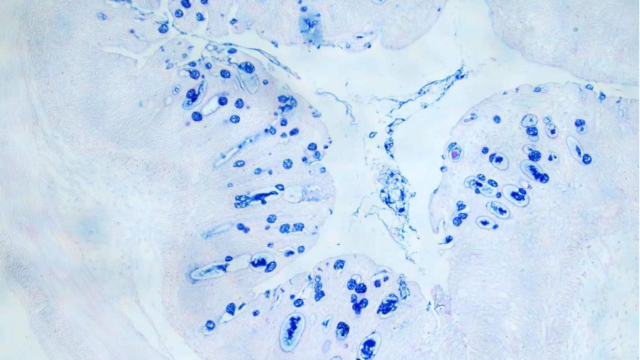
0. 全景扫描在病毒学研究中用于观察病毒的入侵与复制过程,通过高分辨率成像技术捕捉病毒颗粒与宿主细胞表面受体的结合位点、内吞过程及在细胞内的运输路径,其时间分辨率可达毫秒级,能清晰展示病毒脱壳、核酸释放及病毒蛋白合成的动态过程。结合分子生物学技术中的基因编辑、蛋白质印迹等方法,可解析病毒***过程中的关键分子机制,如在研究中,揭示了病毒刺突蛋白与 ACE2 受体结合后的构象变化及病毒进入细胞的具体途径,为抗病毒药物研发提供了病毒***全景动态信息,加速了疫苗和药物的设计进程。贵州免疫组化全景扫描单价
- 河南荧光三标全景扫描单价 2026-01-01
- 新疆荧光全景扫描市场价格 2025-12-31
- 山东髓鞘全景扫描市场价格 2025-12-31
- 山东Masson全景扫描电话多少 2025-12-31
- 山西髓鞘全景扫描性价比 2025-12-31
- 西藏PAS染色全景扫描大概价格 2025-12-30
- 贵州免疫组化全景扫描单价 2025-12-30
- 天津荧光全景扫描大概费用 2025-12-29
- 广西荧光全景扫描大概费用 2025-12-29
- 江苏免疫荧光全景扫描市场价格 2025-12-29
- 山西荧光三标全景扫描大概多少钱 2025-12-26
- 青海髓鞘全景扫描性价比 2025-12-26